
Chasing rabbits: How do soil communities respond to herbivore mammals? Part II
Walter Andriuzzi
Postdoctoral Fellow
Colorado State University
In the previous post I described how herbivore mammals, from rabbits to cattle, can affect the soil food web, and how difficult it is to find general patterns across different places. Here I summarize what we found in a meta-analysis of field studies that had compared soil communities inside and outside exclosures (plots of land that were fenced to keep herbivores out).

Rene van der Wal, ecologist at University of Aberdeen, surveys the plant community on Isle of May. Photo by: Walter Andriuzzi
First, the effects of herbivores depend on climate. For example, soil respiration rate is lower with herbivores than without in subarctic ecosystems, whereas in temperate ecosystems it is higher with herbivores than without. Second, the effects of herbivores vary between trophic groups in the soil food web, and between soil biological functions. For example, unlike respiration, soil microbial biomass and nitrogen mineralization rate do not respond consistently to herbivores in either climate type, whereas they both tend to decline when herbivores are present under arid climates. In subarctic sites, root-feeding nematodes are negatively affected by herbivores, whereas predatory nematodes appear to do just as fine.
Third, the effects of herbivores vary also within trophic groups in the soil food web. Take for example two of the most widespread and abundant groups of soil animal decomposers, oribatid mites and springtails: the former decrease in abundance when herbivores are present, whereas the latter do not show a consistent response. Fourth, the effects of herbivores depend on the herbivore species, in a way that can be at least partly predicted based on their body size: the smaller the herbivore, the more likely its effects on the soil organisms to be neutral, or even positive (that is, increasing biological activity or abundance of soil organisms); the larger the herbivore, the more negative its effects tend to be.
How do our results compare with previous theory? An established framework predicts that herbivore presence has positive effects in the highly productive systems, and negative effects in poorly productive systems. However, the results of our meta-analysis support these predictions only in part. Herbivores have negative effects in low productivity biomes, for example in tundra, but most highly productive systems also exhibited negative effects of herbivory on belowground communities. The big exception was soil respiration in temperate grassland. Moreover, we found that climate is more important than vegetation type; and that herbivore identity and body size are also important.
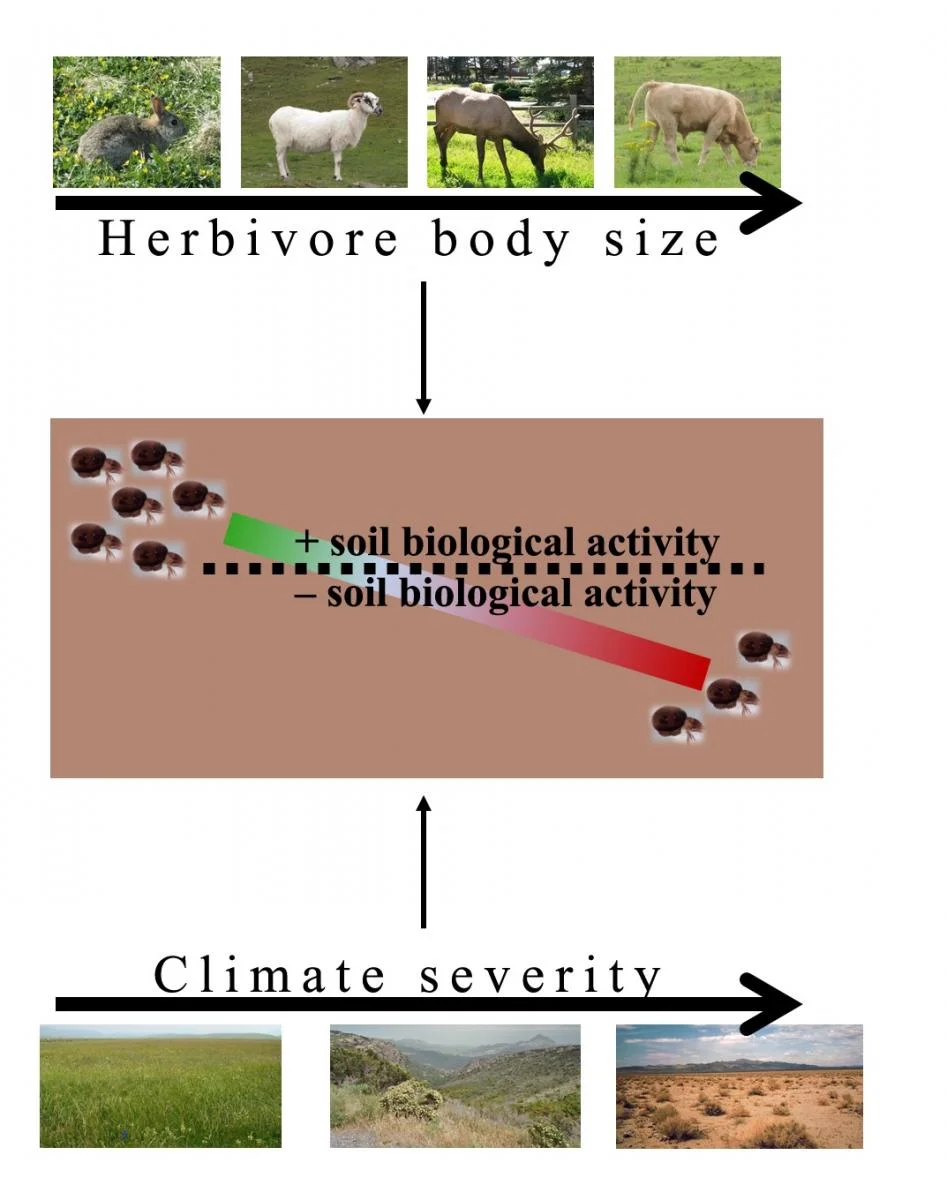
Conceptual model of how herbivore size and climate determine the effect of herbivores on the soil community. Modified from Andriuzzi & Wall 2017
In our meta-analysis, we did observe that responses tend to shift to negative with increasing body size, and an explanation may well be physical disturbance. A more recent framework of belowground responses to herbivory focused on the role of large herbivores on soil compaction. In fine-textured soils, trampling by cows reduces soil pore size and limits oxygen and/or water availability, leading to declines in mineralization rates. We could not test the importance of soil texture in our meta-analysis, because too few of the available studies quantified it rigorously.
Based on our results we developed a conceptual diagram which partly integrates those previous frameworks, by envisioning gradients of herbivore body size and/or climate limitations. Small herbivores, for example the rabbits on the Isle of May, may have limited effects on soil biological activity, or even enhance it by fuelling the soil food web with their excreta and/or more palatable plant resources. Large herbivores, for example cattle (especially if at high density), may lead to physical disturbance such as compaction (because of trampling) and exposure of soil to erosion and atmospheric agents (because of great reduction of plant cover) so the net response belowground is negative. Likewise, with a temperate climate herbivore presence could promote soil biological activity, though not to a great extent if the system is productive enough (as likely was the case on the Isle of May). In ecosystems more limited by aridity, cold, etc., herbivore presence begets the opposite response due to its physical effects on soil. All this is a simplification - different groups of soil organisms may not respond in the same way – but it offers a mechanistic framework to make sense of our findings, and to generate new hypotheses to test.
There are several areas with limited data that challenge the applicability of our general frame work. There were very few studies from the tropics that we could include in the meta-analysis. As a result, while we know a good deal on the effects of reindeer on soil organisms in subarctic tundra, we know next to nothing on those of elephants and impalas in African savannah. Another major surprise was how little some major groups of soil organisms have featured in herbivore exclusion experiments. Protists, which are essential in the soil food web, are lacking altogether from the meta-analysis; earthworms, which have huge effects on soil and plants, were also poorly represented. I hope that our meta-analysis will spur new research to understand how herbivores change their ecosystems not only aboveground, but also belowground.