
Soil animal biogeography? Using oribatid mites as a good start!
Xue Pan, University of Göttingen (Germany)
Every soil biologist has likely asked themself: Why are soil organisms so diverse and abundant even within a small patch of soil; Why have some species a cosmopolitan distribution whereas others are endemic; and is it still possible to collect the same species at the same locations after several decades. How do you answer these questions? I suspect that you would try to explain them from as many perspectives as possible—from species’ morphological characteristics to functional traits, from ancient lineages to present-day communities, and from environmental factors to human activities—in order to make your answers more reasonable and convincing. Congratulations! This means that you are approaching soil biodiversity with a modern biogeographical mindset, a multidisciplinary science dating back to the mid-eighteenth century that attempts to document spatial patterns of biological diversity.
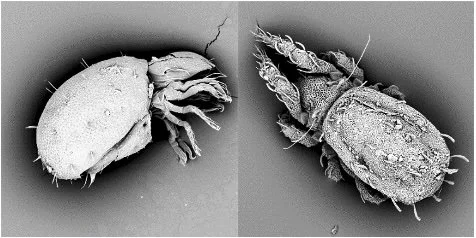
Steganacarus applicatus from Austria (left) and Nothrus anauniensis from China (right) (© Xue Pan, Ulrike Lipka, Mark Maraun)
Soil oribatid mites (Oribatida, Acari) are my favorite study organisms. They are evolutionarily ancient soil animals that originated in the early Paleozoic or possibly even in the Precambrian. They are among the most diverse and abundant belowground animals, with more than 11,000 species described (although up to 100,000 species may exist overall), and they currently occupy virtually all ecosystems.
As an initial attempt in soil Oribatida biogeography, I investigated the distribution of soil oribatid mites at local and regional scales, focusing on the Alps in Europe and Changbai Mountain in the Palearctic region of Asia. I aimed to understand the interplay among historical geological settings, contemporary environmental factors, and species traits in shaping the distribution of soil oribatid mites. Distinct patterns of diversity, community structure, and reproductive modes of soil oribatid mites were observed along altitudinal gradients, with parthenogenesis (i.e., asexual reproduction from unfertilized eggs) dominating among species shared between the two mountain systems. Furthermore, stable isotope analyses (15N; 13C) revealed pronounced differences in trophic niche differentiation, mainly due to differences in parent rock types between the two mountains, thereby elucidating mechanisms underlying oribatid mite species diversity and coexistence in montane soil habitats. Subsequently, by combining phylogenetic and trait-based approaches, we found that soil oribatid mites on Changbai Mountain are phylogenetically older than those in the Alps. Notably, most Changbai Mountain species exhibit broader trophic niches, larger geographical range sizes, and a higher frequency of parthenogenetic reproduction compared with Alpine species. A second attempt was conducted in the Indo-Australian Archipelago, where soil oribatid mite species diversity and endemism were high across all regions studied. Dissimilarity patterns indicate biogeographical regionalization between the Sunda Shelf and the Sahul Shelf, i.e. the Wallace line, which was strongly correlated with geological distance, highlighting the key role of vicariance in shaping oribatid mite distribution patterns.
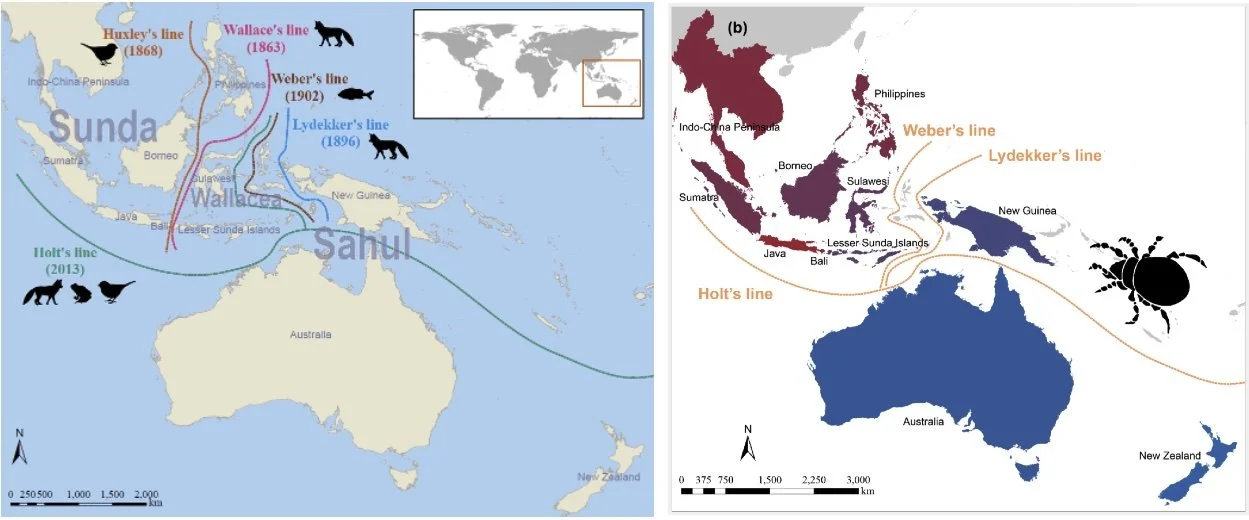
Scheme of the Indo-Australian Archipelago with zoological boundaries based on aboveground and aquatic animal taxa (left) and aboveground taxa indicated by soil oribatid mites (right).
My steps on soil Oribatida biogeography have moved forward to the global scale. However, the first major challenge, also a serious bottleneck, is the lack of credible global distribution data for soil oribatid mites. Another challenge is how to organize published distribution data and, where possible, to incorporate unpublished records. These efforts are represented by the Global Oribatid Occurrence Dataset (GOOD) that I have synthesized to date. Notably, GOOD compiles detailed taxonomic and distributional information on oribatid mites worldwide and continues to grow through the addition of ecological and geographical data. However, GOOD is geographically biased, with the highest data resolution in Europe and the lowest in Africa, where some regions of tropical Africa are nearly devoid of data. Therefore, we advocate increased sampling efforts in underexplored and unexplored regions to reduce data incompleteness and to more accurately reconstruct global soil animal biogeography.
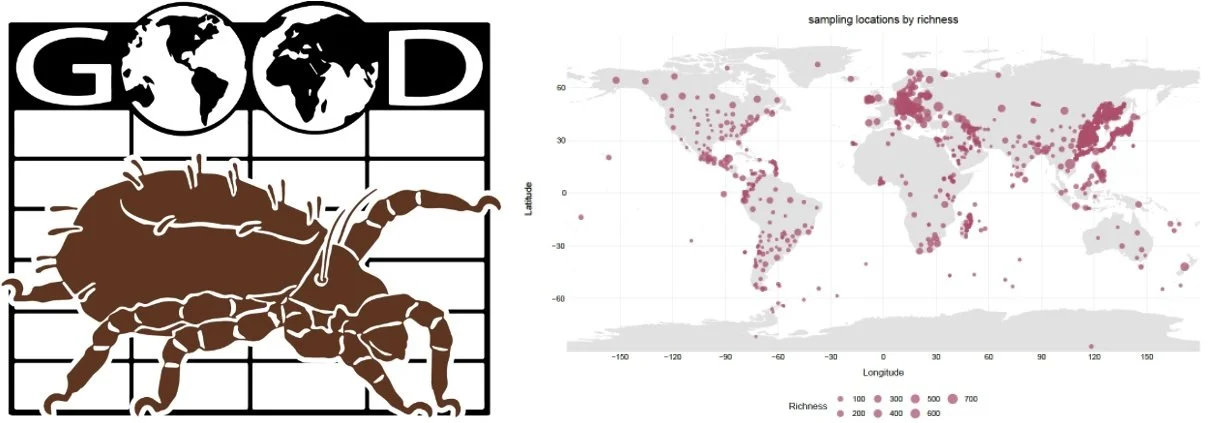
The Global Oribatida Occurrence Dataset (GOOD) and its data coverage (Pan et al., unpublished).